V průběhu zárodečného vývoje nervové trubice vzniká u obratlovců nejen základ centrální nervové soustavy, ale také specifická populace buněk, které následně migrují po těle. Tuto populaci buněk označujeme jako neurální lištu. Z ní je odvozená řada různých buněčných typů, z hlediska počtu řádově srovnatelných s množstvím buněčných typů původem z embryonálních zárodečných listů (ektodermu, entodermu a mesodermu). Z neurální lišty vznikají například odontoblasty tvořící dentin, ale také buňky srdeční svaloviny, chromatofory – buňky zajišťující pigmentaci povrchu těla, nebo mnohé buněčné typy spjaté s periferní nervovou soustavou. Kromě toho však dává neurální lišta vzniknout buněčným typům, které mohou pocházet také z již zmíněných zárodečných listů. Například osteoblasty, buňky tvořící kostní tkáň, mohou pocházet jak z mesodermu, tak z neurální lišty. Předpokládá se, že potenciál diferencovat se v osteoblasty mají přirozeně pouze buňky hlavové neurální lišty, které mohou mj. tvořit většinu kostí lebky. Také odontoblasty vznikají pouze z hlavové neurální lišty.
S tímto stavem se setkáme například u savců, ptáků nebo obojživelníků. Co však další skupiny, různí rybovití obratlovci? Jejich trup často kryjí šupiny či desky původem z odontod, struktur, z nichž vznikly také obratlovčí zuby. Tělní povrch společného předka recentních čelistnatců kryla jak kostěná (osteogenní) složka, tak nad ní ležící „zubovitá“ (odontogenní) složka sestávající z dentinu překrytého sklovinou a podobným materiálem. Na těle dnešních paryb a paprskoploutvých i svaloploutvých ryb pak můžeme nalézt šupiny, štítky či zoubkovité dentikly (též dentikuly) odvozené z tohoto pancíře. Někdy jejich stavba zhruba odpovídá předpokládanému výše popsanému stavu – např. šupiny bichirů sestávají z kosti, dentinu a povrchové vrstvy skloviny (resp. tzv. ganoinu). Naopak dentikly paryb rostoucí na povrchu těla, ale i na méně očekávaných místech, jako je povrch oka či vnitřek žaberních štěrbin, tvoří pouze odontogenní část. U odvozených paprskoploutvých kostnatých ryb (Teleostei) se zase obvykle vyskytují pružné šupiny tvořené jen tenkou vrstvou kosti. To je i případ běžně používaných modelových ryb dánia pruhovaného (Danio rerio) a medaky japonské (Oryzias latipes), jejichž šupiny vznikají pouze z mesodermu.
Před několika lety se ukázalo, že dermální dentikly rejnoka bodlinatého (Leucoraja erinacea) vznikají z odontoblastů trupové neurální lišty. Na to, jaký je podíl trupové neurální lišty na tvorbě šupin paprskoploutvých ryb, si posvítil Jan Štundl se spolupracovníky. Zaměřili se konkrétně na povrchové struktury různých linií paprskoploutvých – bichirů (Cladistia), chrupavčitých (Chondrostei, reprezentovaných jeseterem) a Holostei (reprezentovaných kostlínem) a poté na sumečka krunýřovce (Ancistrus, zástupce Teleostei), u kterého byly šupiny ztraceny a druhotně nahrazeny silnými štítky s dentikly na okraji.
Autoři zjistili, že dentikly bichirů a sumečků vznikají činností odontoblastů odvozených z trupové neurální lišty. Obdobný původ mají také u kostlínů, jejichž šupiny sice postrádají vrstvu dentinu mezi kostí a ganoinem, ve vývoji těchto šupin se však uplatňují drobné dentikly na jejich povrchu. A co osteogeneze, která byla dosud považovaná za schopnost buněk odvozených pouze z hlavové neurální lišty?
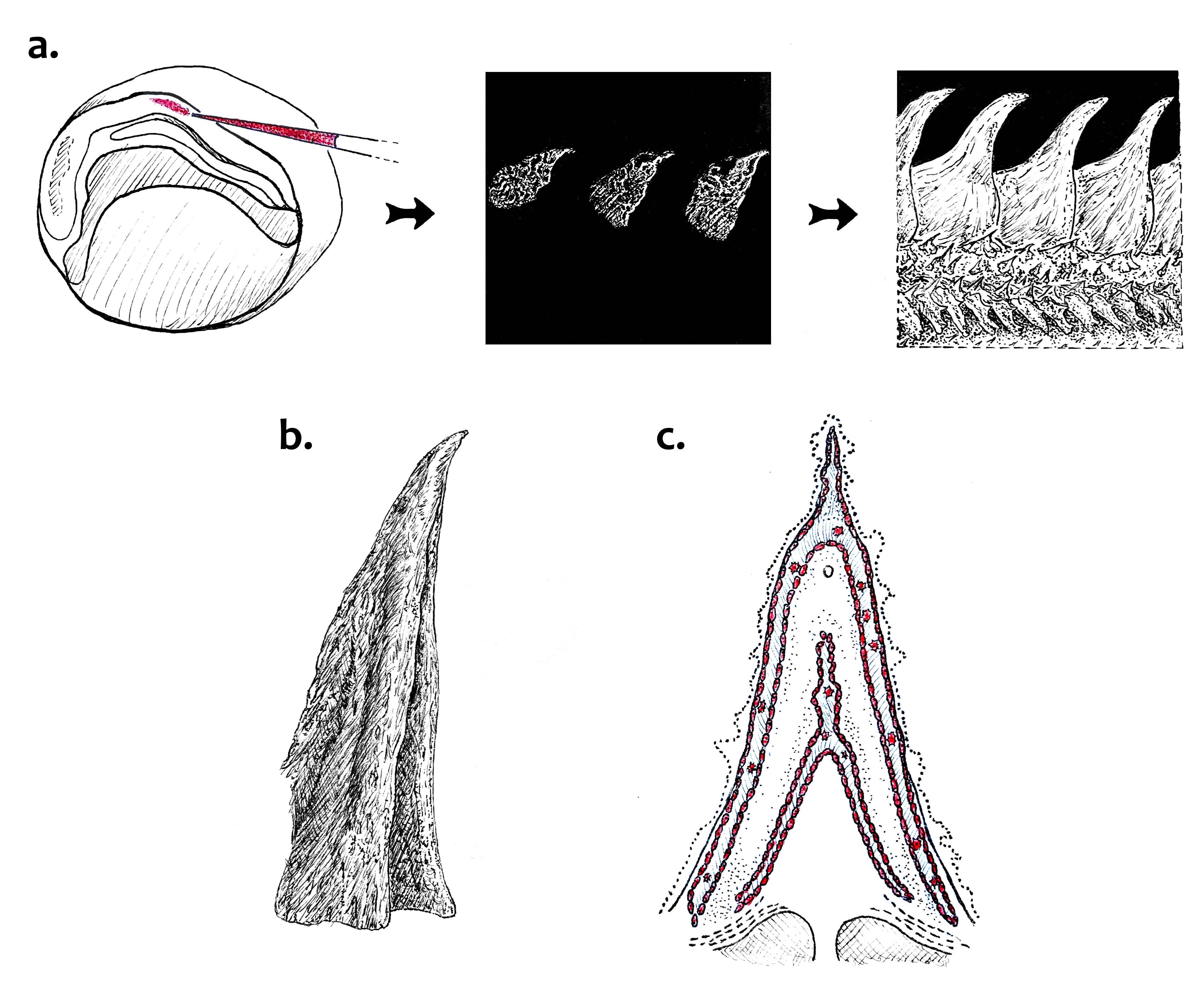
K jejímu studiu se hodí štítky jeseterů, které sestávají pouze z kostní tkáně. I tyto štítky vznikly z původní ganoidní šupiny a zachovaly si pouze kostěnou složku. U embryí jesetera malého (Acipenser ruthenus) proto autoři trasovali migraci buněk trupové části neurální lišty pomocí fluorescenčního barviva. Část těchto buněk se v průběhu vývoje usadila ve vznikajících hřbetních štítcích, a to konkrétně v oblasti, kde se utváří kost. Naopak takto sledované buňky odvozené z paraxiálního mesodermu se na tvorbě kosti štítků nepodílely. Buňky trupové neurální lišty tedy mají osteogenní potenciál alespoň u některých obratlovců. U studovaného jesetera malého a bichira senegalského (Polypterus senegalus) navíc transkriptomické analýzy odhalily expresi genů často spojovaných s genetickým programem neurální lišty. Tyto geny prakticky nebyly exprimovány v mesodermálních šupinách dánia.
Výsledky autorů jsou v souladu s hypotézou, že osteogenní a odontogenní potenciál neurální lišty nebyl vždy omezen pouze na hlavovou neurální lištu. Trupová neurální lišta se tedy mohla podílet na tvorbě kostěných pancířů fosilních obratlovců. Zůstává otázkou, do jaké míry se o tuto funkci dělila s mesodermem. Autoři také spekulují, zda tenké šupiny kostnatých ryb vznikající z mesodermu nemohou představovat novou strukturu, která nahradila původní silné šupiny pocházející z neurální lišty. Srovnávací analýzy zahrnující další obratlovce, zejména svaloploutvé včetně čtyřnožců, kteří nejsou běžně používanými modelovými organismy, jistě pomohou dál vyjasnit potenciál různých částí neurální lišty a například odhalit, jaký je původ dermálního skeletu u některých čtyřnožců – dřívější práce napovídají, že trupová neurální lišta stojí za vznikem osteodermů pásovců nebo dermálních kostí plastronu želvího krunýře.